Simulation results & figures
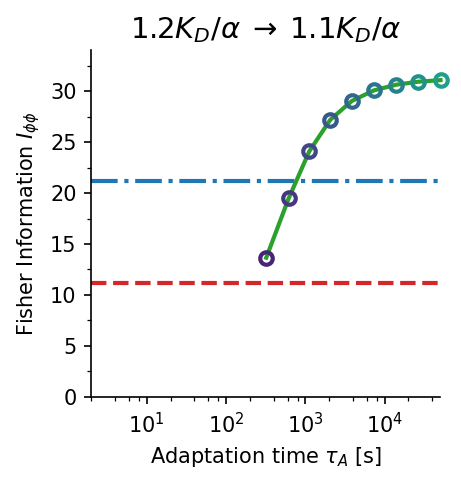
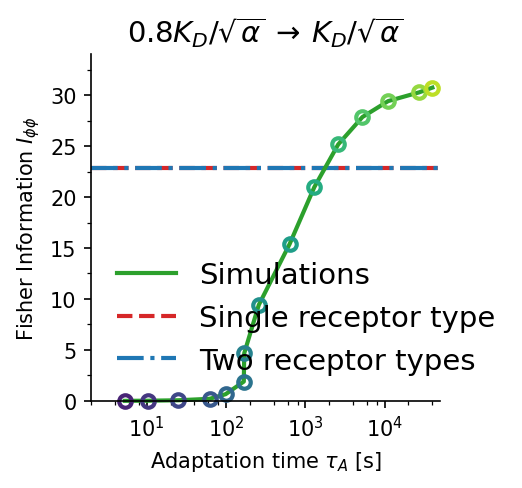
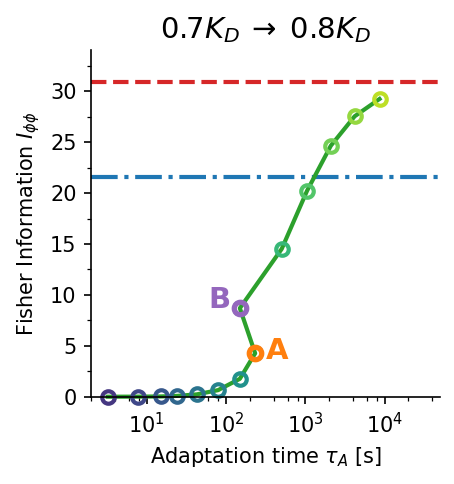
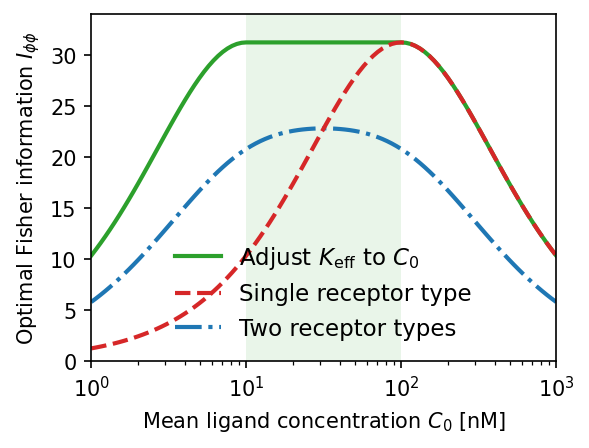
Fisher information comparison across receptor models. The allosteric adaptation model (green) maintains near-maximum $I_{\phi\phi}$ over the full range $C_0 \in [K_D/\alpha, K_D]$ (shaded). Single receptor type (red dashed) peaks sharply at $K_D$. Two receptor types (blue dash-dot) broaden the range but sacrifice peak accuracy. $K_D = 100\,\text{nM}$, $\alpha = 10$.
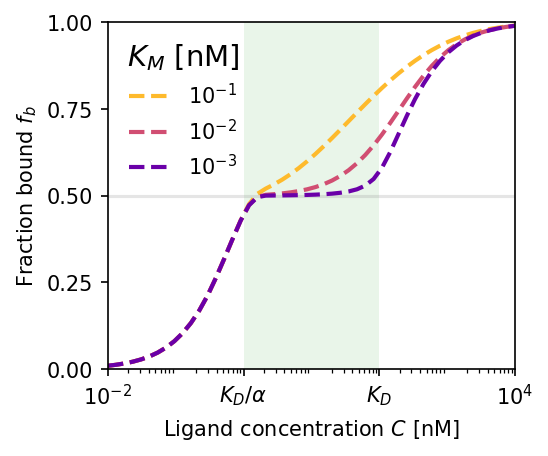
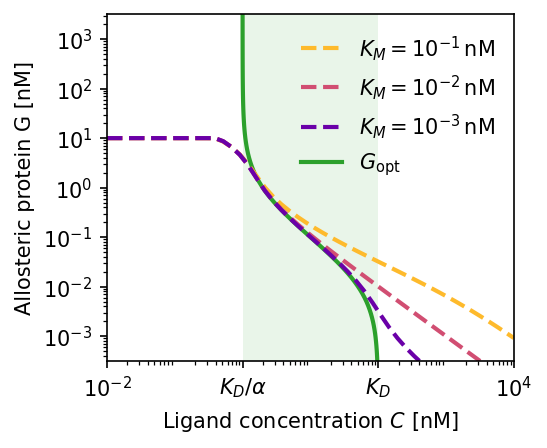
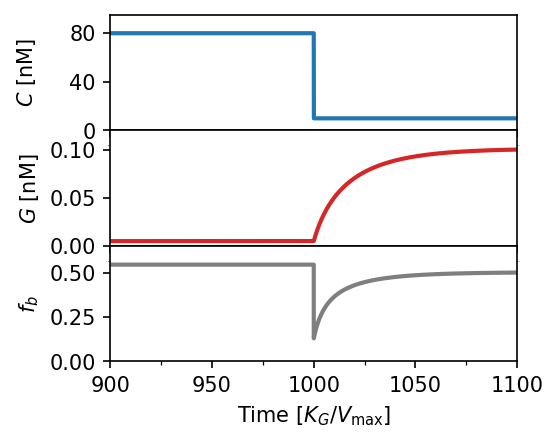
Adaptation dynamics. (a) Steady-state bound fraction $f_b$ vs. ligand concentration. Smaller Michaelis constant $K_M$ yields better adaptation over a wider concentration range (green region). (b) Corresponding active allosteric protein concentration $G$; green line is the theoretical optimum. (c) Time-series response after a sudden drop $80 \to 10\,\text{nM}$: $G$ adapts over ~$100\,t$ units, restoring $f_b \approx 1/2$. $K_D = 100\,\text{nM}$, $\alpha = 100$, $K_M = 10^{-3}\,\text{nM}$.
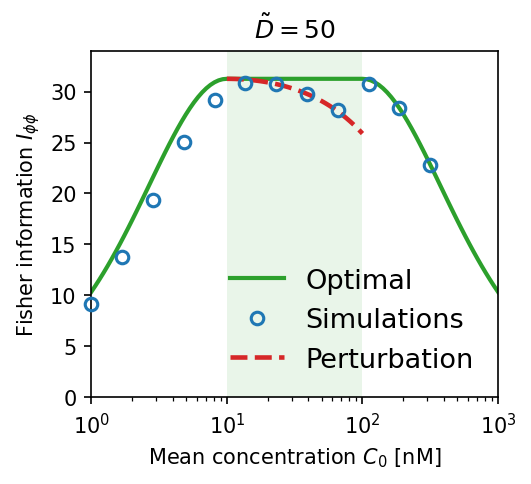
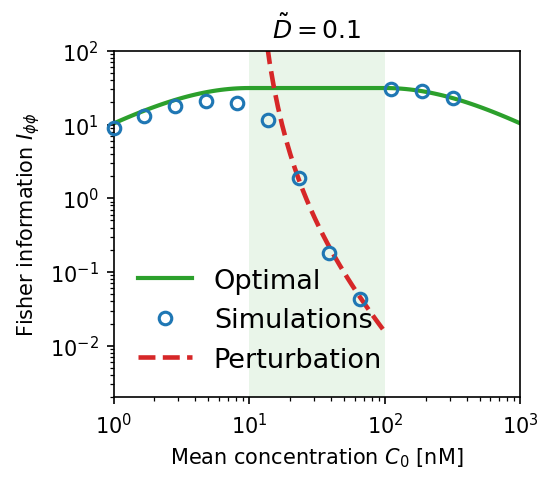
Effect of allosteric protein diffusivity. Blue circles are numerical simulations; red dashed lines are first-order perturbation theory; solid green is the ideal (optimal) limit. (a) High diffusion $\tilde{D} = 50$: simulations match theory and approach optimal $I_{\phi\phi}$. (b) Low diffusion $\tilde{D} = 0.1$: Fisher information collapses by orders of magnitude — local adaptation destroys the gradient signal.
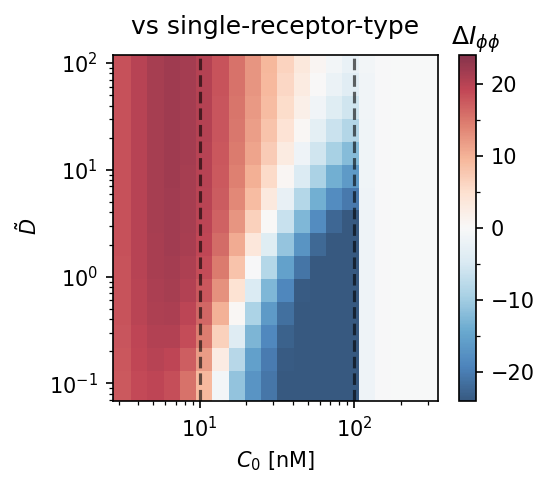
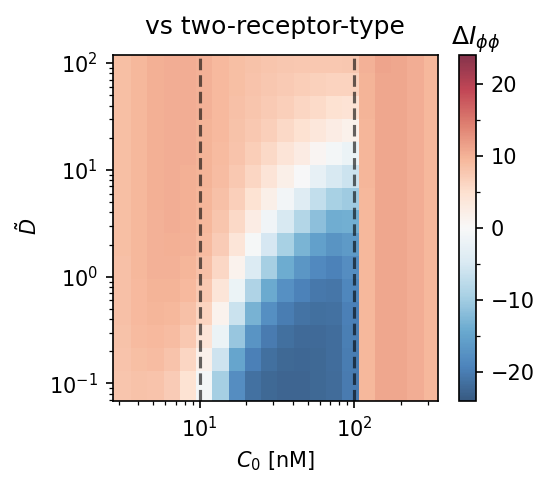
Model comparison heatmaps. $\Delta I_{\phi\phi} = I_{\phi\phi}^\text{adapt} - I_{\phi\phi}^\text{baseline}$ as a function of diffusivity $\tilde{D}$ and mean concentration $C_0$. Red = adaptation wins. The adaptation model requires $\tilde{D} \gtrsim 10$–$100$ to outperform single-receptor type (left) or two-receptor-type (right) strategies. Dashed lines: $C_0 = K_D/\alpha$ and $K_D$.
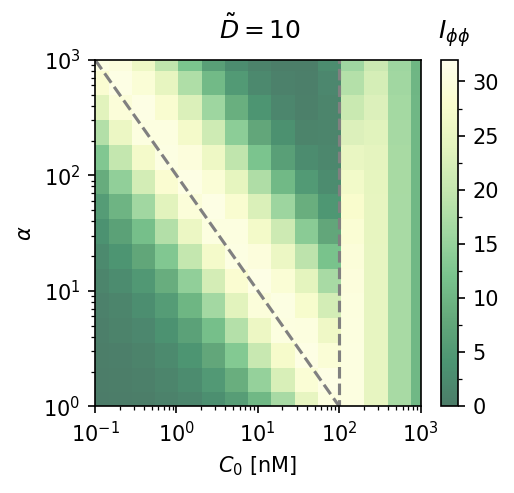
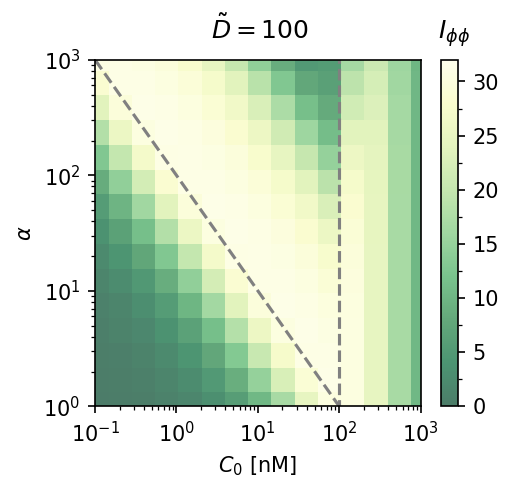
Cooperativity $\alpha$ vs. mean concentration. Fisher information heatmaps at $\tilde{D} = 10$ (left) and $\tilde{D} = 100$ (right). Larger $\alpha$ extends the accurate sensing range only at high $\tilde{D}$; at moderate diffusion, increasing $\alpha$ mainly shifts the region of accuracy.
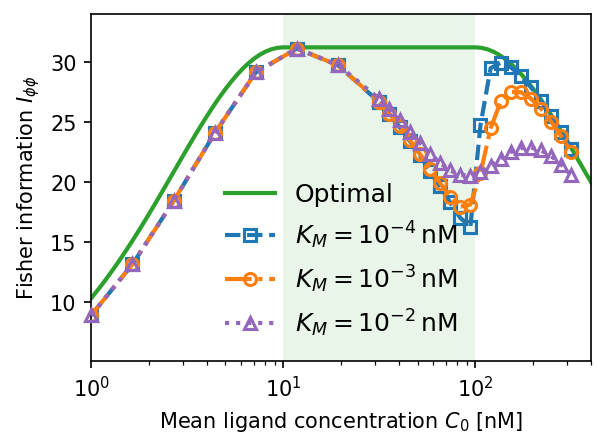
Dependence on Michaelis constant $K_M$. Smaller $K_M$ sharpens the jump in Fisher information at $C_0 = K_D$ (near-perfect adaptation for $C_0 \in [K_D/\alpha, K_D]$). Larger $K_M$ smooths the transition and reduces peak accuracy. $\tilde{D} = 10$, $K_D = 100\,\text{nM}$, $\alpha = 10$.
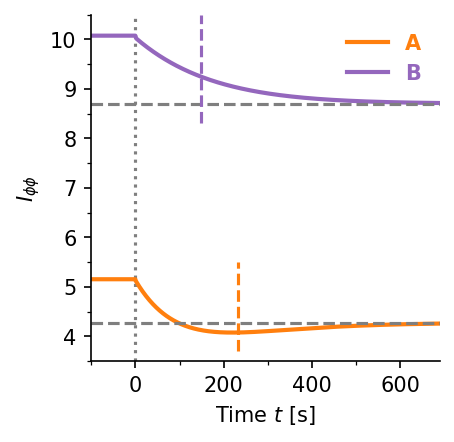
Accuracy–time trade-off. Gradient sensing accuracy $I_{\phi\phi}$ vs. adaptation time $\tau_A$ for three concentration jump scenarios (a–c). Higher Fisher information requires longer adaptation times ($\tau_A \sim 100$–$1000\,\text{s}$). Panel (d) shows the time evolution of $I_{\phi\phi}(t)$ for two representative cases, illustrating the non-monotone dynamics. Blue dash-dot: two-receptor-type baseline; red dashed: single-receptor-type baseline.